物質簡介
DNA由脫氧核苷酸組成的大分子聚合物。脫氧核苷酸由堿基、脫氧核糖和磷酸構成。其中堿基有4種:腺嘌呤(A)、鳥嘌呤(G)、胸腺嘧啶(T)和胞嘧啶(C)。
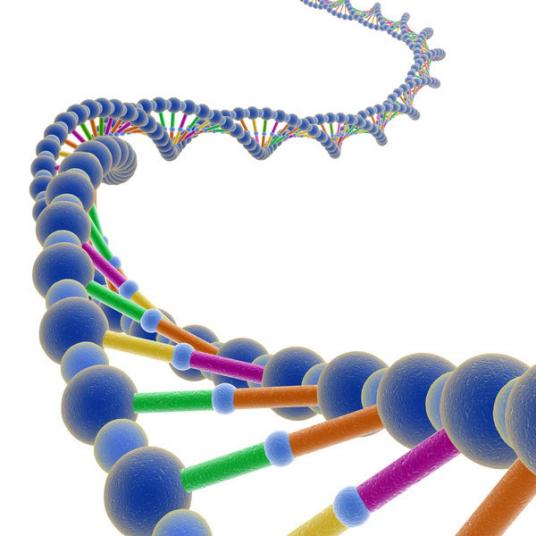
DNA 分子結構中,兩條多脫氧核苷酸鍊圍繞一個共同的中心軸盤繞,構成雙螺旋結構。脫氧核糖-磷酸鍊在螺旋結構的外面,堿基朝向裡面。兩條多脫氧核苷酸鍊反向互補,通過堿基間的氫鍵形成的堿基配對相連,形成相當穩定的組合。
脫氧核糖核酸(DNA)是生物細胞内攜帶有合成RNA和蛋白質所必需的遺傳信息的一種核酸,是生物體發育和正常運作必不可少的生物大分子。
DNA中的核苷酸中堿基的排列順序構成了遺傳信息。該遺傳信息可以通過轉錄過程形成RNA,然後其中的mRNA通過翻譯産生多肽,形成蛋白質。
在細胞分裂之前,DNA複制過程複制了遺傳信息,這避免了在不同細胞世代之間的轉變中遺傳信息的丢失。 在真核生物中,DNA存在于細胞核内稱為染色體的結構中。在沒有細胞核的其它生物中,DNA要麼存在于染色體中要麼存在于其它組織(細菌有單環雙鍊DNA分子,而病毒有DNA或RNA基因組)。在染色體中,染色質蛋白如組蛋白、共存蛋白和凝聚蛋白将DNA在一個有序的結構中。這些結構指導遺傳密碼和負責轉錄的蛋白質之間的相互作用,有助于控制基因的轉錄。
曆史
DNA最初是由瑞士生物化學家弗裡德裡希·米歇爾(Friedrich Miescher)1869年從手術繃帶的膿液中分離出來的,由于這種微觀物質位于細胞核中,當時被稱為核蛋白(nuclein)。
1919年,Phoebus Levene确定了DNA由含氮堿基,糖和磷酸鹽組成的核苷酸結成。Levene提出DNA由一條通過磷酸鹽結合在一起的核苷酸組成。他确信DNA長鍊較短,且其中的堿基是以固定順序重複排列。
1937年,William Astbury展示了第一個X射線衍射研究的結果,表明DNA具有極其規則的結構。
1928年,英國科學家弗雷德裡克·格裡菲斯(1877-1941)在實驗中發現,平滑型的肺炎球菌,能轉變成為粗糙型的同種細菌。 該系統在沒有提供任何物質引起變化的證據的同時,表明某些物質可以将遺傳信息從死亡細菌的遺體傳遞給生物。1943年奧斯瓦爾德·埃弗裡等人的試驗證明DNA是這一轉變現象背後的原因。
1944年,Erwin Schrödinger鑒于量子物理學少數原子的系統具有無序行為理論,斷言遺傳物質必須由大的非重複分子構成,方足以維持遺傳信息的穩定。
1953年由Alfred Hershey和Martha Chase通過另一個經典實驗得到證實DNA在遺傳中的作用最終在,該實驗表明噬菌體T2的遺傳物質實際上是DNA,而蛋白質則是由 DNA的指令合成的。
1953年,美國的沃森和英國的克裡克提出了DNA雙螺旋結構的分子模型。
1958年,馬修·梅瑟生與富蘭克林·史達在梅瑟生-史達實驗中,确認了DNA的複制機制。後來克裡克團隊的研究顯示,遺傳密碼是由三個堿基以不重複的方式所組成,稱為密碼子。
1961年,哈爾·葛賓·科拉納、羅伯特·W·霍利及馬歇爾·沃倫·尼倫伯格解出這些密碼子所構成的遺傳密碼。
組成
DNA是由重複的核苷酸單元組成的長聚合物,鍊寬2.2到2.6納米,每個核苷酸單體長度為0.33納米。盡管每個單體占據相當小的空間,但DNA聚合物的長度可以非常長,因為每個鍊可以有數百萬個核苷酸。例如,最大的人類染色體(1号染色體)含有近2.5億個堿基對。
生物體中的DNA幾乎從不作為單鍊存在,而是作為一對彼此緊密相關的雙鍊,彼此交織在一起形成一個叫做雙螺旋的結構。每個核苷酸由可與相鄰核苷酸共價鍵結合的側鍊骨架和含氮堿基組成,兩條鍊上的含氮堿基通過堿基互補以氫鍵相連。糖與含氮堿基形成核苷,核苷與一個或多個磷酸基團結合成為核苷酸。
DNA骨架結構是由磷酸與糖類基團交互排列而成。組成脫氧核糖核酸的糖類分子為環狀的2-脫氧核糖,屬于五碳糖的一種。磷酸基團上的兩個氧原子分别接在五碳糖的3号及5号碳原子上,形成磷酸雙酯鍵。這種兩側不對稱的共價鍵位置,使每一條脫氧核糖核酸長鍊皆具方向性。雙螺旋中的兩股核苷酸互以相反方向排列,這種排列方式稱為反平行。脫氧核糖核酸鍊上互不對稱的兩末端一邊叫做5'端,另一邊則稱3'端。脫氧核糖核酸與RNA最主要的差異之一,在于組成糖分子的不同,DNA為2-脫氧核糖,RNA則為核糖。
DNA的雙螺旋通過在兩條鍊上存在的含氮堿基之間建立的氫鍵來穩定。組成DNA的四種堿基是腺嘌呤(A)、胞嘧啶(C)、鳥嘌呤(G)和胸腺嘧啶(T)。所有四種堿基都具有雜環結構,但結構上腺嘌呤和鳥嘌呤是嘌呤的衍生物,稱為嘌呤堿基,而胞嘧啶和胸腺嘧啶與嘧啶有關,稱為嘧啶堿基。
兩條核苷酸鍊沿着中心軸以相反方向相互纏繞在一起,很像一座螺旋形的樓梯,兩側扶手是兩條多核苷酸鍊的糖一磷基因交替結合的骨架,而踏闆就是堿基。DNA雙螺旋是右旋螺旋。不同磷酸鹽基團之間的凹槽仍然暴露在外。主溝寬2.2納米,而小溝寬1.2納米。兩個凹槽的不同寬度決定了蛋白質對不同堿基的可接觸性,這取決于堿基是在主溝還是小溝中。與DNA的蛋白質,如轉錄因子,通常與處在大溝中的堿基接觸。
結構
一級結構
DNA的一級結構,是指4種核苷酸的連接及其排列順序,表示了該DNA分子的化學構成。
DNA的一級結構決定其高級結構,如B-DNA中多G-C區易形成左手螺旋DNA(Z-DNA),而反向重複的DNA片段易出現發夾結構等。這些高級結構又決定和影響着一級結構的功能。
二級結構
DNA的二級結構是指兩條多核苷酸鍊反向平行盤繞所生成的雙螺旋結構。
通常情況下,DNA的二級結構分兩大類:一類是右手螺旋,如A-DNA和B-DNA,DNA通常是以右手螺旋形式存在的;另一類是左手螺旋,即Z-DNA。
不同螺旋形式DNA分子主要參數比較
高級結構
DNA的高級結構,是指DNA雙螺旋進一步扭曲盤繞所形成的更複雜的特定空間結構,包括超螺旋、線性雙鍊中的紐結(kink)、多重螺旋等。其中,超螺旋結構是DNA高級結構的主要形式,可分為正超螺旋(右手超螺旋)與負超螺旋(左手超螺旋)兩大類,負超螺旋是細胞内常見的DNA高級結構形式,正超螺旋是過度纏繞的雙螺旋。它們在不同類型的拓撲異構酶作用下或在特殊情況下可以相互轉變。
理化性質
DNA是高分子聚合物,其溶液為高分子溶液,具有很高的粘度,可被甲基綠染成綠色。DNA對紫外線(260nm)有吸收作用,利用這一特性,可以對DNA進行含量測定。當核酸變性時,吸光度升高,稱為增色效應;當變性核酸重新複性時,吸光度又會恢複到原來的水平。較高溫度、有機溶劑、酸堿試劑、尿素、酰胺等都可以引起DNA分子變性,即DNA雙鍊堿基間的氫鍵斷裂,雙螺旋結構解開—也稱為DNA的解螺旋。
主要類别
單鍊DNA
單鍊DNA(single-stranded DNA)大部分DNA以雙螺旋結構存在,但一經熱或堿處理就會變為單鍊狀态。單鍊DNA就是指以這種狀态存在的DNA。單鍊DNA在分子流體力學性質、吸收光譜、堿基反應性質等方面都和雙鍊DNA不同。某些噬菌體粒子内含有單鍊環狀的DNA,這樣的噬菌體DNA在細胞内增殖時則形成雙鍊DNA。
閉環DNA
閉環DNA(closed circular DNA)沒有斷口的雙鍊環狀DNA,亦稱為超螺旋DNA。由于具有螺旋結構的雙鍊各自閉合,結果使整個DNA分子進一步旋曲而形成三級結構。另外如果一條或二條鍊的不同部位上産生一個斷口,就會成為無旋曲的開環DNA分子。從細胞中提取出來的質粒或病毒DNA都含有閉環和開環這二種分子。可根據兩者與色素結合能力的不同,而将兩者分離開來。
垃圾DNA
垃圾DNA(Junk DNA)是指生物體内不翻譯成蛋白質的DNA,過去多認為它們無用,所以稱為垃圾DNA。後來,科學家發現垃圾DNA中包含有重要的調節機制,從而能夠控制基礎的生物化學反應和發育進程,這将幫助生物進化出更為複雜的機體。生物越複雜,垃圾DNA似乎就越重要。
生物功能
在基因組中,遺傳信息存儲在稱為基因的DNA序列中,這個遺傳信息的傳遞由互補的含氮堿基序列的存在得到保證。事實上,在轉錄過程中,遺傳信息可以很容易地被轉錄到互補的RNA鍊中(mRNA)。mRNA通過翻譯合成蛋白質。或者,細胞可以通過稱為DNA複制的過程簡單地複制遺傳信息。
基因組結構
真核生物基因組DNA位于細胞核内,線粒體和葉綠體内也有DNA。原核生物DNA被包裹在細胞質中不含細胞膜的不規則細胞器類核中。 遺傳信息包含在基因中,基因是能夠影響生物體表型的遺傳單位。每個基因含有開放閱讀框(能夠轉錄成RNA的區域)和由啟動子和增強子組成的調節區。 在許多物種中,隻有一小部分基因組序列可以被轉錄和翻譯。例如,人類基因組中隻有1.5%序列含有編碼蛋白質的外顯子,超過50%的人類基因組由重複的非編碼DNA序列組成。在任何情況下,不編碼蛋白質的DNA序列也可以轉錄成非編碼RNA,參與基因表達的調控。 一些非編碼序列是對染色體的結構組成部分。端粒和着絲粒區域通常含有非常少的基因,但對于染色體的功能和穩定性是必需的。
轉錄和翻譯
基因是含有能夠影響生物體表型特征的遺傳信息的DNA序列。基因内的DNA堿基序列作為模闆可以合成RNA分子,在大多數情況下,RNA分子被翻譯成多肽,最終稱為蛋白質。 将基因的核苷酸序列複制到RNA鍊中的過程稱為轉錄,由RNA聚合酶催化發生。 RNA鍊有不同的命運:一些RNA分子實際上具有結構(例如在核糖體内發現的那些rRNA)或催化(如核酶)功能;絕大多數RNA經曆成熟過程産生mRNA,被翻譯成蛋白質。 翻譯過程發生在細胞質中,其中mRNA與核糖體結合,并由遺傳密碼介導。核糖體允許順序讀取mRNA密碼子,有利于它們識别和與特定tRNA相互作用,這些tRNA攜帶對應于每個單個密碼子的氨基酸分子。
遺傳密碼
遺傳密碼是一組規則,将DNA或RNA序列以三個核苷酸為一組的密碼子轉譯為蛋白質的氨基酸序列,以用于蛋白質合成。密碼子由mRNA上的三個核苷酸(例如ACU,CAG,UUU)的序列組成,每三個核苷酸與特定氨基酸相關。例如,三個重複的胸腺嘧啶(UUU)編碼苯丙氨酸。使用三個字母,可以擁有多達64種不同的組合。由于有64種可能的三聯體和僅20種氨基酸,因此認為遺傳密碼是多餘的(或簡并的):一些氨基酸确實可以由幾種不同的三聯體編碼。但每個三聯體将對應于單個氨基酸。最後,有三個三聯體不編碼任何氨基酸,它們代表停止(或無意義)密碼子,分别是UAA,UGA和UAG 。
DNA複制
DNA複制是指DNA雙鍊在細胞分裂以前進行的複制過程,複制的結果是一條雙鍊變成兩條一樣的雙鍊(如果複制過程正常的話),每條雙鍊都與原來的雙鍊一樣。這個過程是通過名為半保留複制的機制來得以順利完成的。複制可以分為以下幾個階段:
起始階段:解旋酶在局部展開雙螺旋結構的DNA分子為單鍊,引物酶辨認起始位點,以解開的一段DNA為模闆,按照5'到3'方向合成RNA短鍊。形成RNA引物。
DNA片段的生成:在引物提供了3'-OH末端的基礎上,DNA聚合酶催化DNA的兩條鍊同時進行複制過程,由于複制過程隻能由5'→3'方向合成,因此一條鍊能夠連續合成,另一條鍊分段合成,其中每一段短鍊成為岡崎片段(Okazaki fragments)。
RNA引物的水解:當DNA合成一定長度後,DNA聚合酶水解RNA引物,補填缺口。
DNA連接酶将DNA片段連接起來,形成完整的DNA分子。
最後DNA新合成的片段在旋轉酶的幫助下重新形成螺旋狀。
與蛋白質作用
所有DNA功能都取決于其與特定蛋白質的相互作用。這些相互作用可以是非特異性的,也可以是極其特異性的。還有許多可以結合DNA的酶,其中,在DNA轉錄和複制中複制DNA序列的聚合酶特别重要。
DNA與組織蛋白(圖1中白色部分)的交互作用,這種蛋白質中
的堿性氨基酸(左下藍色),可與DNA上的酸性磷酸基團結合(右下紅色)。
結構蛋白可與DNA結合,是非專一性DNA-蛋白質交互作用的常見例子。染色體中的結構蛋白與DNA組合成複合物,使DNA組織成緊密結實的染色質構造。對真核生物來說,染色質是由脫DNA與一種稱為組織蛋白的小型堿性蛋白質所組合而成;而原核生物體内的此種結構,則摻雜了多種類型的蛋白質。
DNA可在組織蛋白的表面上附着并纏繞整整兩圈,以形成一種稱為核小體的盤狀複合物。組織蛋白裡的堿性殘基,與DNA的酸性糖磷酸骨架之間可形成離子鍵,使兩者發生非專一性交互作用,也使複合物中的堿基序列相互分離。
在堿性氨基酸殘基上所發生的化學修飾有甲基化、磷酸化與乙酰化等,這些化學作用可使DNA與組織蛋白之間的作用強度發生變化,進而使DNA與轉錄因子接觸的難易度改變,影響轉錄作用的速率。其他位于染色體内的非專一性DNA結合蛋白,還包括一種能優先與DNA結合,并使其扭曲的高移動性群蛋白。這類蛋白質可以改變核小體的排列方式,産生更複雜的染色質結構。
DNA結合蛋白中有一種專門與單鍊DNA結合的類型,稱為單鍊DNA結合蛋白。人類的複制蛋白A是此類蛋白中獲得較多研究的成員,作用于多數與解開雙螺旋有關的過程,包括DNA複制、重組以及DNA修複。這類結合蛋白可固定單鍊DNA,使其變得較為穩定,以避免形成莖環(stem-loop),或是因為核酸酶的作用而水解。
相對而言,其他的蛋白質則隻能與特定的DNA序列進行專一性結合。大多數關于此類蛋白質的研究集中于各種可調控轉錄作用的轉錄因子。這類蛋白質中的每一種,都能與特定的DNA序列結合,進而活化或抑制位于啟動子附近序列的基因轉錄作用。轉錄因子有兩種作用方式,第一種可以直接或經由其他中介蛋白質的作用,而與負責轉錄的RNA聚合酶結合,再使聚合酶與啟動子結合,并開啟轉錄作用。第二種則與專門修飾組織蛋白的酵素結合于啟動子上,使DNA模闆與聚合酶發生接觸的難度改變。
由于目标DNA可能散布在生物體中的整個基因組中,因此改變一種轉錄因子的活性可能會影響許多基因的運作。這些轉錄因子也因此經常成為信号傳遞過程中的作用目标,也就是作為細胞反映環境改變,或是進行分化和發育時的媒介。具專一性的轉錄因子會與DNA發生交互作用,使DNA堿基的周圍産生許多接觸點,讓其他蛋白質得以“讀取”這些DNA序列。多數的堿基交互作用發生在大凹槽,也就是最容易從外界接觸堿基的部位。
結合DNA的酶
核酸酶和連接酶:核酸酶是能夠切割DNA鍊的酶,因為它們催化磷酸二酯鍵的水解。從位于DNA鍊末端的核苷酸開始水解DNA的核酸酶稱為核酸外切酶。另一方面,直接切入DNA鍊的那些是内切核酸酶。分子生物學中使用最廣泛的核酸酶,稱為限制性内切酶,以切割特定序列的DNA。在自然界中,這種酶通過在進入細菌細胞時消化噬菌體DNA來保護細菌免受噬菌體感染。通常,限制性核酸酶識别特定的回文核苷酸序列,稱為限制性位點。這些酶廣泛用于涉及在載體内亞克隆DNA的技術中。
DNA連接酶:是能夠使用來自ATP或NAD的化學能将先前切割或斷裂的DNA鍊聚集在一起的酶。連接酶在DNA滞後鍊複制中特别重要,因為它們将岡崎碎片組合成DNA鍊。連接酶在DNA修複和基因重組中也發揮重要作用。
拓撲異構酶和解旋酶: 拓撲異構酶是具有活性核酸酶和連接酶的酶。這些酶能夠改變DNA的拓撲特性。它們中的一些通過切割DNA螺旋并允許其旋轉,降低其超螺旋程度,然後通過連接酶将兩端連接。另一方面,其它拓撲異構酶能夠在連接斷裂的DNA鍊之前,切斷螺旋,并允許第二個螺旋通過斷裂部位。拓撲異構酶是許多涉及DNA的過程所必需的,例如DNA複制和轉錄。解旋酶是能夠利用核苷三磷酸中存在的化學能的蛋白質,尤其是ATP,以破壞核堿基之間形成的氫鍵,從而允許DNA的雙螺旋打開成單鍊。
聚合酶:聚合酶是從核苷三磷酸合成多核苷酸鍊的酶。它們通過向鍊上存在的先前核苷酸的3'-OH添加核苷酸起作用。因此,所有聚合酶都以5' - 3'方向起作用。DNA複制需要DNA依賴的DNA聚合酶,實現DNA序列的完美拷貝。有些DNA聚合酶具有校對功能,能夠檢測含氮堿基之間的錯配錯誤并激活3'或5'外切核酸酶作用以去除不正确的堿基。 在大多數生物體中,DNA聚合酶在稱為replisoma的較大蛋白質複合物中起作用,該複合體由許多酶例如解旋酶組成。 RNA依賴的DNA聚合酶是使用RNA片段作為模闆合成DNA的特殊類聚合酶,包括逆轉錄酶(一種參與逆轉錄病毒感染的病毒酶)和端粒酶(它是端粒複制所必需的)。 與DNA依賴性DNA聚合酶一樣,這些RNA依賴的DNA聚合酶也在由輔助分子和調節分子組成的廣泛蛋白質複合物中起作用。
應用領域
法醫鑒定
通常從血液、皮膚、唾液、頭發和其它組織和體液中分離DNA,以識别罪犯或犯罪行為。常用的遺傳指紋識别。該技術比較重複DNA的可變區段的長度,例如短串聯重複序列和小衛星,它們在個體之間有不同。因此,檢查中的兩個DNA樣品之間的比較不是基于對整個DNA序列的分析,而是僅基于這些重複序列部分。事實上,兩個沒有血緣關系的個體間99.9%的DNA序列是相同的。這種方法通常非常可靠,但犯罪現場被其他人的DNA污染時,對罪犯的識别會很複雜。 這種方法由英國遺傳學家Sir Alec Jeffreys于1984年開發。遺傳指紋識别也可用于識别群體性事件的受害者。 未經同意采集DNA的行為稱為基因盜竊。
基因工程
現代生物學和生物化學大量使用DNA。術語重組DNA是指人工構建和組裝的DNA片段。它們可以以質粒的形式或通過其它類型的載體整合插入到生物體中。由此産生的生物被稱為轉基因生物。可用于生産重組蛋白,用于生物醫學研究或農業栽培。